
Les membres de l’Afas publient régulièrement des articles. Ils sont à retrouver ici :
Jeanne Brugère-Picoux
Professeur honoraire de pathologie médicale du bétail et des animaux de basse-cour (Ecole nationale vétérinaire d’Alfort), membre de l’Académie nationale de médecine, présidente honoraire de l’Académie vétérinaire de France

L'étude «Une enquête sérologique à grande échelle chez les animaux de compagnie d'octobre 2020 à juin 2021 en France montre une exposition significativement plus élevée au Sras-CoV-2 chez les chats» [1] correspond à un projet de recherche ANR CoVet. Elle a été conduite et dirigée par trois vétérinaires, dont deux appartenant à l’Académie nationale de médecine (Eric Leroy et Serge Rosolen) et la troisième à l’Académie vétérinaire de France (Alexandra Briend-Marchal).
Il s’agit de la même équipe qui avait publié dans One Health [2] une enquête réalisée avec le CHRU de Besançon regroupant médecins, vétérinaires et chercheurs. Ils avaient alors démontré, dès 2020, l’importance d’une contamination des animaux par leur propriétaires infectés. Ils avaient aussi noté la plus forte prévalence de l’infection par le Sars-CoV-2 chez le chat (vingt chats positifs sur trente-quatre, soit 58,8%) par comparaison avec le chien (cinq chiens séropositifs sur treize, soit 38,5%).
Enfin signalons que deux des auteurs qui ont piloté cette enquête sont membres actifs de la veille informationnelle mise à jour deux fois par mois sur le site de l’Académie nationale de médecine.
De nombreuses études avaient montré, après les deux cas observés chez des chiens à Hong Kong en 2020, que l’agent de la Covid-19 (Sars-CoV-2) pouvait infecter de nombreux animaux. Ce fut surtout le cas des animaux de compagnie. Cependant, il s’agissait le plus souvent d’un nombre limité d’animaux étudiés et ce, pendant une période limitée. Ceci peut expliquer la disparité des résultats obtenus : de 0 à 14,5% chez les chiens et de 0 à 21,7% chez les chats.
Une vaste enquête sérologique réalisée d’octobre 2020 à juin 2021 (pendant les deuxième et troisième vagues de l’épidémie) chez 2036 chats et 3577 chiens prélevés lors de consultations en clinique vétérinaire a permis une évaluation plus précise de la séroprévalence en France de l’infection par le Sars-CoV-2.
Cette étude a permis de noter que les chats étaient plus souvent séropositifs que les chiens (9,3% et 5,9% respectivement). Rappelons qu'à la différence des chiens, les chats sont plus sensibles à l’infection par le Sras-CoV-2 et qu’ils peuvent se transmettre la maladie.
Comparée aux résultats observés en Europe (de 0 à 6,4%) la plus forte prévalence chez les chats français dans cette étude peut s’expliquer par le fait que les prélèvements européens ont eu lieu principalement lors de la première vague, où le virus circulait moins. Cependant il n’y a pas eu une augmentation de la séroprévalence au cours des huit mois de l’étude comme les auteurs pouvaient le penser avec la persistance des anticorps dans l’organisme. Cela pourrait s’expliquer par la persistance limitée des anticorps (deux études réalisées chez sept chiens et quatre chats infectés naturellement ont montré une persistance des anticorps variant de trois à dix mois). Si c’est le cas, cette étude sérologique réalisée sur huit mois ne refléterait pas la proportion réelle de chats infectés, avec une sous-estimation du nombre réel de chats infectés
Par ailleurs cette étude a permis de noter qu’il n’y avait pas de différence selon le sexe des animaux prélevés. Au contraire, les auteurs ont pu noter une séroprévalence plus faible chez les animaux plus âgés. Cette plus forte séroprévalence chez les jeunes animaux âgés de moins de trois ans peut s’expliquer par un plus important contact avec leur propriétaire à cette période mais aussi à une diminution plus rapide des taux d’anticorps chez l’animal âgé comme cela a été observé chez l’Homme.
Cette étude souligne l'importance d'une approche en «santé globale» face à la pandémie de Covid-19 et pose la question de la vaccination des animaux de compagnie en contact étroit avec l'Homme. En effet, on ne peut exclure un risque de recombinaison virale chez l’animal avec un risque de transmission en retour du virus vers l’Homme (rappelons la récente transmission du Sars-CoV-2 d’un chat à une vétérinaire en Thaïlande).
Bien que les animaux de compagnie ne semblent pas actuellement jouer un rôle dans la pandémie en cours, ces résultats soulignent que l'ampleur de l'infection par le Sras-CoV-2 chez les animaux de compagnie n'est pas anodine. En raison de l’importante population de chats et de chiens domestiques en contact étroit avec leurs propriétaires, les auteurs soulignent l'importance de collecter davantage de données sur la transmissibilité et la pathogénicité du Sars-CoV-2 chez les animaux de compagnie, notamment avec la surveillance des souches virales isolées (RT-qPCR, suivi par séquençage du génome entier pour identifier une éventuelle mutation). Il importe aussi de rappeler les recommandations en santé publique à mettre en œuvre pour permettre de prévenir la transmission de l'Homme.
[1] Fritz M, Elguero E, Becquart P, de Riols de Fonclare D, Garcia D, Beurlet S, Denolly S, Boson B, Rosolen SG, Cosset FL, Briend-Marchal A, Legros V, Leroy EM. A large-scale serological survey in pets from October 2020 through June 2021 in France shows significantly higher exposure to SARS-CoV-2 in cats (bioRxiv preprint doi: https://doi.org/10.1101/2022.12.23.521567), 26 décembre 2022.
[2] Fritz M, Rosolen B, Kraft E, E, Becquart P, Elguero E, Vratskikh O, Denolly S, Boson B, Vanhomwegen J, Ar Gouilh M, Kodjo A, Chirouze C, Rosolen SG, Legros V, Leroy EM. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households, One Health, 11 (2020), 100192 (https://doi.org/10.1016/j.onehlt.2020.100192)
Alain Delacroix
Professeur honoraire, chaire «Chimie industrielle - Génie des procédés» du Conservatoire national des arts et métiers

Le 26 septembre 1887, au congrès de l’Association française pour l’avancement des sciences, un «électricien, ingénieur à Paris» présente plusieurs appareils liés à la fée électricité : des polyscopes cliniques, une lampe de sécurité, un photophore, une lampe sous-marine et un nouvel interrupteur. Mais ce qui étonne le plus les participants, c’est l’utilisation de l’auxanoscope électrique quand ils voient projetés sur un écran les portraits de Chevreuil, de Pasteur et de divers objets.
Qui est ce Géo Trouvetout français ? Il s’agit de Gustave Trouvé qui, avec l’initiale de son prénom, a un patronyme prédestiné – G. (j'ai) trouvé ! Né en 1839 à La Haye-Descartes en Indre-et-Loire, il est le fils d’un marchand de bétail. Après des études au collège de Chinon, il apprend la serrurerie et commence en 1855 des études à l’Ecole des arts et métiers d’Angers, qu’il ne terminera pas. Il part alors à Paris et travaille chez un horloger. A partir de 1865, il habite 5 rue Montesquieu et y produit son premier brevet : une pile minuscule portative permettant l’animation des bijoux et divers objets d’art. En 1866, il déménage au 6 rue Thérèse et c’est là qu’il enregistre sa marque de fabrique Eureka – «j'ai trouvé» en grec ancien, ce qui correspond à l’initiale de son prénom et à son nom. Il va alors produire un nombre considérable d’inventions. Il aide, entre autres, Léon Foucault à résoudre le problème de la durée de rotation de son gyroscope en y installant un système électro-moteur. C'est Henri Tresca, sous-directeur au Conservatoire des arts et métiers, qui a suggéré à Foucault de rencontrer Gustave Trouvé, de vingt ans son cadet.
En 1867, il expose un fusil électrique à l’Exposition universelle de Paris, qui est très apprécié par l’empereur Napoléon III. En 1869, il présente un explorateur extracteur électrique de corps étrangers ayant pénétré dans les tissus organiques, qui sera très utile lors de la guerre de 1870. En 1873, il produit le polyscope qui sert à explorer le corps humain.
Le 1er juillet 1878, il s’installe au 14 rue Vivienne et y installe son atelier et y poursuit ses inventions : un moteur électrique, un tricycle électrique qui est le premier véhicule de ce type au monde, un bateau à propulsion électrique, une machine à coudre électrique, le polyscope et le photophore qui servent d’éclairage électrique pour l’usage des médecins mais aussi pour les spectacles, une lampe de sécurité, l’auxanoscope qui est l’ancêtre des projecteurs, etc.
En 1902, en travaillant sur un appareil utilisant la lumière ultraviolette pour traiter les maladies de peau, il se blesse, sa plaie s’infecte et il meurt de septicémie à l’hôpital Saint-Louis le 27 juillet. Gustave Trouvé n’ayant pas de descendants et ses archives à la mairie de La Haye-Descartes ayant brûlé en 1980, il tombe dans l’oubli. Même ses restes seront jetés dans la fosse commune lorsque la concession de sa tombe sera arrivée à échéance.
Pourtant on doit à Gustave Trouvé plus de soixante-dix inventions liées à l’électricité. Grâce à un historien anglais, Kevin Desmond, qui a écrit sa biographie, on commence à le réhabiliter et plusieurs plaques ont été déposées aux endroits où il a habité. Le musée national des techniques du Conservatoire national des Arts et Métiers possède dix-neuf objets construits par Gustave Trouvé, mais à ce jour, aucun n’est exposé.
Alain Delacroix
Professeur honoraire, chaire "Chimie industrielle - Génie des procédés" du Conservatoire national des arts et métiers

Au XIXe siècle, la banane est pratiquement inconnue en France car son transport par bateau est trop long et les fruits arrivent inconsommables. C’est seulement en 1900 que l’on commence à voir des bananes à la vente à Paris. Le propriétaire d’une bananeraie à Madère fait venir les fruits verts et les fait mûrir dans une mûrisserie boulevard de Rochechouart. Moyennant quoi les bananes sont très chères. Néanmoins la consommation de ce fruit augmente rapidement à Paris et en 1904, une société anglo-américaine, Fyffes, capte le marché. Cette société existe encore en 2022 et est la plus vieille entreprise liée au commerce de la banane. Après les années trente, la Martinique et la Guadeloupe vont largement augmenter leur production et en 1939, la flotte française de bananier était la quatrième flotte mondiale.
Depuis le début, le transport des bananes depuis des régions lointaines posait des problèmes car les fruits mûrissaient de façon intempestive au cours d’un voyage trop long. Elles étaient transportées vertes dans des cales autour de 12 °C avec ventilation, puis on les réchauffait dans des mûrisseries. Le chauffage était réalisé par combustion du gaz de ville. Il semblerait qu’un jour, un propriétaire de mûrisserie moderniste a voulu chauffer avec l’électricité et que les bananes ne mûrissaient plus correctement. Cela semblait prouver que ce n'était pas la chaleur uniquement qui provoquait le mûrissement des fruits.
Indépendamment de cela, on s’était rendu compte au XIXe siècle que les arbres situés à côté de lampadaires à gaz perdaient leurs feuilles plus vite que les autres. En 1901, on s’est aperçu que les semis de pois noirs présentent d’étranges symptômes quand ils sont cultivés dans une salle utilisant du gaz de charbon. En 1910, on a observé que les émanations d’oranges provoquent la maturation accélérée des bananes et enfin en 1934, on a découvert que l’éthylène est une hormone végétale. A partir de ce moment, on s’est rendu compte que c’était la très faible concentration en éthylène liée à la combustion du gaz de ville qui favorisait le mûrissement des bananes.
L’éthylène se révélant être une hormone végétale, on a voulu en trouver de nombreuses applications sur les cultures. Le problème est que l’éthylène est un gaz qui, agissant en faible concentration, ne peut être utilisé dans les champs. On a donc recherché un produit liquide ou soluble dans l’eau, susceptible de se décomposer en éthylène. Celui qui a été retenu est l’éthéphon, Cl-CH2-CH2-PO3H2, qui se décompose en éthylène, acide chlorhydrique et acide phosphorique (chlorures et phosphates). Outre l’éthylène, ses produits de décomposition aux concentrations utilisées sont inoffensifs. Quant à l’éthéphon lui-même, il a une très faible toxicité avec une LD 50 (dose létale médiane) de l’ordre de 3000 mg/kg pour le rat. Toutefois il faut prendre des précautions particulières quand il est concentré. L’éthéphon est présent dans de très nombreux produits commerciaux dont l’Ethrel. En tant que régulateur de croissance des plantes, il est très utilisé. On peut citer quelques effets :
- il prévient la verse des céréales,
- c’est une substance de croissance du riz, du blé, du coton, du café, des bananes, du tabac, etc.,
- il favorise l’éclaircissage des pommes et favorise la formation des bourgeons floraux pour l’année suivante.
Ce produit, très utilisé et aux multiples avantages, vient d’être cité dans les médias et est à l’origine d’un probable «scandale» dont ceux-ci sont friands. L’éthéphon aurait été utilisé dans des bananeraies pour faire jaunir des bananes plantain alors qu’il est interdit en Martinique dans la culture des bananes. En revanche, il est autorisé dans la culture des ananas. On en a alors profité pour le relier aux problèmes liés au chlordécone. Ces deux molécules n’ont pourtant aucun rapport. L’une est un régulateur de croissance des plantes, soluble dans l’eau et rapidement dégradée, alors que l’autre est un pesticide donc biocide, très peu soluble dans l’eau et très stable donc persistant dans l’environnement.
Il est dommage qu’une molécule aussi utile ne soit décrite dans les médias qu’à l’occasion d’un éventuel «scandale», sans donner d’informations sur ses bienfaits. Cela participe à la méfiance du public envers la chimie, et les sciences en général, ce qui n’est pas très rassurant pour l’avenir.
Jeanne Brugère-Picoux
Professeur honoraire de pathologie médicale du bétail et des animaux de basse-cour (Ecole nationale vétérinaire d’Alfort), membre de l’Académie nationale de médecine, présidente honoraire de l’Académie vétérinaire de France

Un article paru dans The Lancet le 10 août 2022 rapporte l’observation en France d’un premier cas mondial de transmission du virus de la variole du singe de l’Homme au chien [1]. Le chien contaminé appartenait à deux hommes ayant consulté à l’hôpital de la Pitié-Salpêtrière le 10 juin et présentant les lésions classiques de la maladie signalées depuis début mai dans leur communauté homosexuelle : asthénie, céphalées, hyperthermie, ulcérations anales et, chez l’un d’eux, éruption vésiculo-pustuleuse sur le visage, les oreilles et les jambes. L’infection virale a été confirmée par PCR sur les prélèvements réalisés chez les deux patients (peau et oropharynx pour l’un, anus et oropharynx pour l’autre). Douze jours après l’apparition de ces symptômes, leur chien lévrier mâle âgé de quatre ans, sans antécédent de maladie, a présenté des lésions cutanéo-muqueuses (pustules sur l’abdomen et fine ulcération anale). Par PCR, le virus de la variole du chien a été identifié sur divers prélèvements (pustules cutanées, anus, cavité buccale). Il était identique à celui des propriétaires (et au clade hMPXV-1, lignée B.1 répandu depuis avril 2022 dans les régions non endémiques). Les propriétaires ont signalé qu’ils dormaient avec le chien dans leur chambre tout en ayant évité tout contact de ce chien avec d’autres animaux ou des personnes dès l’apparition de leurs premiers symptômes (soit 13 jours avant les troubles cutanés chez le chien).
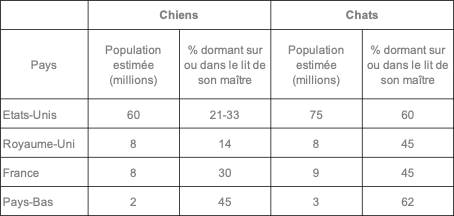
Le risque lié à la présence d’un chien ou d’un chat dans la chambre de son maître n’est pas nouveau. Il peut même paraître surprenant de constater le nombre de personnes acceptant de laisser leur animal dormir dans leur lit. Une enquête publiée en 2011 et réalisée de 1974 à 2010 aux Etats-Unis, aux Pays-Bas, en France et au Royaume-Uni a permis de noter que 14 à 62% des propriétaires dorment avec leur chat ou leur chien (cf. tableau). On oublie combien cette habitude peut favoriser la transmission de diverses maladies (bartonellose ou maladie des griffes du chat ; Capnocytophaga canimorsus, bactérie présente dans la salive du chien et pouvant provoquer une septicémie mortelle ; pasteurellose ; staphylococcies ; parasitoses) [2]. Dans le cas de la Covid-19, une enquête canadienne a montré que le temps passé avec un contact entre le chien et son maître n’avait pas joué un rôle dans la contamination de l’animal mais qu’il n’en était pas de même pour les chats qui étaient plus à risque d’être contaminés s’ils dormaient dans le lit de leur maître [3].
Dans cette enquête, les poxvirus n’étaient pas cités. Alors que la variole a été la première maladie pouvant être prévenue par la vaccination et ainsi la première à être éradiquée, les autres infections à poxvirus ont souvent été sous-estimées alors que certaines sont zoonotiques. En premier lieu, il faut souligner que ces poxviroses zoonotiques sont souvent bénignes, sauf chez les personnes immunodéprimées. C’est pourquoi l’infection humaine reste une zoonose sous-estimée car peu fréquente et de ce fait rarement reconnue immédiatement par le clinicien (cela a pu être aussi le cas pour la variole du singe, qui a pu ainsi se propager pendant un certain temps sans être détectée avant mai 2022 en dehors de l’Afrique du fait de son mode particulier de propagation par des relations sexuelles dans la communauté homosexuelle masculine). Les poxvirus les plus souvent rencontrés chez l’Homme sont le cowpox (orthopoxvirus) et l’ecthyma contagieux (parapoxvirus), très connu chez les petits ruminants mais beaucoup moins des particuliers propriétaires de moutons ou de chèvres «tondeuses». Dans le cas du cowpox, des cas graves exceptionnels ont cependant pu être observés. Bien que l’infection par le cowpox soit sporadique, le virus est endémique en Europe du Nord, avec une augmentation du nombre de cas depuis une vingtaine d’années, peut-être du fait de l’arrêt de la vaccination contre la variole [4]. Le réservoir de ce poxvirus est constitué par des petits rongeurs sauvages, notamment le rat d’égout ou surmulot (Rattus norvegicus), espèce dont dérive le rat domestique d’élevage, facile à apprivoiser comme rat de laboratoire ou nouvel animal de compagnie (NAC). C’est ainsi qu’en Europe nous avons connu en 2009 une épidémie humaine avec des rats importés de Hongrie en tant que NAC [5]. Le rat peut infecter l’Homme et de nombreuses espèces animales, le plus souvent par morsure (chiens, chats, bovins, chevaux, singes, lamas, éléphants…). Si les cas sont rares chez le chien (infecté par morsure de la truffe), ils sont plus fréquents chez le chat du fait de son comportement de chasseur (le premier cas félin de cowpox a été décrit en 1978). Ce contact étroit du chat avec les rongeurs explique qu’il soit la source la plus importante des infections humaines.
Dans le cas de l’orthopoxvirus de la variole du singe, en dehors des singes utilisés dans des laboratoires à partir desquels le virus fut isolé pour la première fois en 1958 (d’où la dénomination de ce virus), on ne connaît pas l’espèce hôte réservoir de ce virus. On a souvent considéré en République démocratique du Congo (RDC), où la maladie est endémique, que l’infection humaine résultait d'un contact avec un animal sylvestre infecté, bien que l’espèce hôte réservoir soit actuellement inconnue. Diverses espèces sont suspectées, qu’il s’agisse d’animaux sauvages devenus familiers ou chassés pour leur consommation (viande de brousse) : le Cricetomys gambianus (rat de Gambie), le Cercopithecus ascanius (singe à queue rousse) et les écureuils africains, en particulier les genres Funisciurus et Heliosciurus rufobrachium (écureuil soleil à pattes rouges) [6,7]. Le seul MPXV isolé d'un mammifère sauvage a été obtenu à partir d'un écureuil à cordes moribond (Funisciurus anerythrus) collecté lors d'une enquête sur une épidémie en RDC [8,9].
L’exemple de l’épidémie de variole du singe observée en mai 2003 dans le Midwest américain est significatif pour plusieurs raisons :
- elle a confirmé l’origine zoonotique suspectée en Afrique de cette poxvirose par l’importation du Ghana de rats de Gambie (ou cricétomes des savanes) [10], ces rats africains d’origine sauvage apparemment sains étant vendus comme nouveaux animaux de compagnie (NAC) ;
- ces rats de Gambie ont joué le rôle de réservoir asymptomatique comme dans le cowpox avec le rat d’égout (ou de compagnie) en contaminant dans l’animalerie des chiens de prairie (Cynomys ludovicianus), autres rongeurs NAC autochtones de la famille des Sciuridae ;
- les chiens de prairie furent malades mais aussi les vecteurs secondaires d’une contamination humaine avec 71 cas, dont plusieurs enfants ;
- les huit premiers cas humains n’ont pas été reconnus (comme ce fut le cas pour les premiers patients atteints par la nouvelle épidémie de variole du singe qui présentaient des lésions génitales) et il a fallu plus d’une semaine pour que ces cas soient signalés aux autorités de santé publique [11] ;
- ce fut la seule importante épidémie de variole simienne observée dans un pays non africain et due à des animaux de compagnie.
La contamination d’un chien par ses propriétaires faisant partie de la communauté homosexuelle, contaminée dans plus de 95% des cas observés depuis mai 2022* dans la pandémie actuelle, ajoute une nouvelle espèce sensible au MPXV. Cet agent pathogène réémergent ne se limite plus aux régions endémiques africaines et présente le risque mondial d’occuper la niche écologique laissée vacante par la variole. Le problème sera de savoir si le MPXV s’établira plus dans un réservoir animal que dans la population humaine s’il continue à se propager.
Connaissant le grand nombre d’espèces animales sensibles à un autre orthopoxvirus zoonotique (cowpox), on ne peut exclure dans ce cas la possibilité d’autres espèces sensibles au MPXV, en particulier parmi les animaux de compagnie, et le risque de rongeurs porteurs asymptomatiques. C’est pourquoi nous soulignerons à nouveau la recommandation de l’Académie nationale de médecine, dans son communiqué du 8 juillet 2022, «d’éviter le contact entre les cas et les animaux pendant la maladie jusqu’à la chute des croûtes» [13]. Une épidémiosurveillance concernant le risque d’une contamination zoonotique par le MPXV de l’Homme vers l’animal ou de l’animal vers l’Homme et l’application stricte des mesures de biosécurité préconisées pendant la période d’isolement de 21 jours dans les cas humains s’avère nécessaire.
En conclusion, nous ne pouvons que reprendre celle de notre confrère Alexis Lécu, responsable du zoo de Vincennes, à propos du Sars-CoV-2 [14] : «Les tribulations de ce virus chez l’animal nous enseignent finalement la même leçon que son apparition en 2019 : nous devons rapidement remettre de la distance avec l’animal, pour notre bien à tous».
[1] Seang S, Burrel S, Todesco E, Leducq V, Monsel G, Le Pluart D, et al. Evidence of human-to-dog transmission of monkeypox virus. The Lancet. août 2022;S0140673622014878.
[2] Chomel BB, Sun B. Zoonoses in the Bedroom. Emerg Infect Dis. févr 2011;17(2):167‑72.
[3] Bienzle D. Cats may catch Covid-19 from sleeping on their owner’s bed. EurekAlert. European Congress of Clinical Microbiology & Infectious Diseases (ECCMID, 30-Jun-2021); 2021.
[4] Rosone F, Sala MG, Cardeti G, Rombolà P, Cittadini M, Carnio A, et al. Sero-Epidemiological Survey of Orthopoxvirus in Stray Cats and in Different Domestic, Wild and Exotic Animal Species of Central Italy. Viruses. 19 oct 2021;13(10):2105.
[5] Campe H, Zimmermann P, Glos K, Bayer M, Bergemann H, Dreweck C, et al. Cowpox Virus Transmission from Pet Rats to Humans, Germany. Emerg Infect Dis. mai 2009;15(5):777‑80.
[6] Walker M. Monkeypox Virus Hosts and Transmission Routes: A Systematic Review of a Zoonotic Pathogen. Biological Sciences Undergraduate Honors Theses [Internet]. 1 mai 2022; Disponible sur: https://scholarworks.uark.edu/biscuht/69
[7] Monroe BP, Doty JB, Moses C, Ibata S, Reynolds M, Carroll D. Collection and Utilization of Animal Carcasses Associated with zoonotic Disease in Tshuapa District, the Democratic Republic of the Congo, 2012. Journal of Wildlife Diseases. juill 2015;51(3):734‑8.
[8] Reynolds MG, Carroll DS, Olson VA et al. C. A Silent Enzootic of an Orthopoxvirus in Ghana, West Africa: Evidence for Multi-Species Involvement in the Absence of Widespread Human Disease. The American Journal of Tropical Medicine and Hygiene. 1 avr 2010;82(4):746‑54.
[9] Jezek Z, Grab B, Szczeniowski MV, Paluku KM, Mutombo M. Human monkeypox: secondary attack rates. Bull World Health Organ. 1988;66(4):465‑70.
[10] Centers for Disease Control and Prevention (CDC). Update: multistate outbreak of monkeypox--Illinois, Indiana, Kansas, Missouri, Ohio, and Wisconsin, 2003. MMWR Morb Mortal Wkly Rep. 11 juill 2003;52(27):642‑6.
[11] Reed KD, Melski JW, Graham MB, Regnery RL, Sotir MJ, Wegner MV, et al. The Detection of Monkeypox in Humans in the Western Hemisphere. N Engl J Med. 22 janv 2004;350(4):342‑50.
[12] CDC. 2022 Monkeypox Outbreak Global Map [Internet]. 2022 [cité 16 août 2022]. Disponible sur: https://www.cdc.gov/poxvirus/monkeypox/response/2022/world-map.html.
[13] Académie nationale de médecine. Variole du singe: zoonose et infection sexuellement transmissible (IST) [Internet]. Communiqué; 2022. Disponible sur: https://www.academie-medecine.fr/variole-du-singe-zoonose-et-infection-sexuellement-transmissible-ist/.
[14] Lécu A. Covid : retour sur 30 mois de tribulations d’un virus humain chez l’animal [Internet]. The Conversation. [cité 27 juill 2022]. Disponible sur: http://theconversation.com/covid-retour-sur-30-mois-de-tribulations-dun-virus-humain-chez-lanimal-187552
Jeanne Brugère-Picoux
Professeur honoraire de pathologie médicale du bétail et des animaux de basse-cour (Ecole nationale vétérinaire d’Alfort), membre de l’Académie nationale de médecine, présidente honoraire de l’Académie vétérinaire de France

La situation évoluant rapidement, le présent article actualise le précédent, daté du 23 mai 2022.
Introduction
On assiste, depuis le 7 mai 2022, à une émergence inhabituelle de cas humains sporadiques de la variole du singe – maladie endémique jusqu’alors localisée à l’Afrique centrale et occidentale – dans plusieurs pays européens, en Amérique du Nord et en Australie. La particularité de ces cas est qu’ils concernent des personnes n’ayant pas voyagé en Afrique, à l’exception du premier malade au Royaume-Uni, qui revenait du Nigéria. Contrairement à la plupart des cas africains, pour lesquels on reconnaît une origine zoonotique, ces cas émergents sont liés à des contaminations interhumaines, souvent observées chez des hommes homosexuels ou bisexuels avec plusieurs partenaires.
Il s’agit aussi d’une zoonose et de nombreux réservoirs animaux sont suspectés en Afrique. C’est pourquoi l’apparition de cette maladie due à un virus très résistant dans le milieu extérieur en dehors de l’Afrique doit amener à une certaine prudence pour éviter tout risque d’instauration d’un réservoir animal autochtone dans ces nouveaux pays touchés par la variole du singe.
Historique
Le virus de la variole du singe a été découvert en 1958 dans un laboratoire de Copenhague chez des macaques (Macaca Fascularis) qui avaient été importés de Singapour [1]. Il y eut deux foyers à quatre mois d’intervalle. Les reins de ces singes servaient à la préparation de cultures cellulaires pour la production de vaccins contre la poliomyélite. C’est pourquoi ce nouveau virus fut dénommé Monkeypox virus (ou MPXV). D’autres laboratoires ont connu ce même problème aux Etats-Unis, aux Pays-Bas et en France [2, 3]. Il s’agissait toujours de singes captifs provenant de diverses régions tropicales (Inde, Malaisie, Philippines et Sierra Leone). Ces cas rapportés chez des singes n’ont jamais donné lieu à une contamination humaine.
Virus de la variole du singe
Il s’agit d’un Orthopoxvirus, gros virus à ADN de la famille des Poxviridae.
Il faut souligner la forte résistance dans le milieu extérieur de ce virus, en particulier à la fin de la maladie dans les croûtes cutanées.
Le terme de monkeypox pourrait changer selon l’Organisation mondiale de la Santé (OMS), qui ne souhaite plus attribuer aux virus émergents un nom discriminatoire ou stigmatisant pouvant évoquer une région géographique ou un animal. D’ailleurs on peut noter une certaine confusion lorsque l’on parle de chickenpox qui est en fait la varicelle, de smallpox pour la variole ou de cowpox pour le virus de la vaccine.
Deux souches virales sont connues pour le MPXV : un clade présent en Afrique centrale (souche Congo), le plus virulent, pouvant provoquer une mortalité de 10,6%, et un clade d’Afrique occidentale, moins pathogène avec un taux de mortalité estimé de 3,6% [4].
Une analyse métagénomique réalisée au Portugal [5] sur des prélèvements réalisés dans plusieurs pays confirme que le virus appartient au clade de l’Afrique occidentale et qu’il semble reconnaître une origine unique qui serait une souche exportée du Nigéria en 2018 et 2019.
Variole du singe chez l’Homme
Au cours des années suivant la première identification d’un cas humain en Afrique – en 1970 en République démocratique du Congo (RDC) chez un enfant de 9 mois –, la variole du singe, qui n’était plus masquée par la variole ou la vaccination, a été régulièrement observée en Afrique centrale et occidentale. Elle est devenue l’infection à orthopoxvirus la plus répandue chez l’Homme [7].
L’âge médian des malades africains est passé de 4 ans dans les années soixante-dix à 21 ans depuis 2010 [4] du fait du déclin de l'immunité collective à la suite de l'arrêt de la vaccination contre la variole qui permettait d’offrir une protection croisée estimée à 85% [9].
Lors de l’épidémie de variole du singe en Afrique de l'Ouest qui a débuté en septembre 2017 au Nigéria, quatre personnes voyageant du Nigéria vers le Royaume-Uni [10], Israël [11] et Singapour [12] sont devenues les premiers cas humains exportés d'Afrique. Il y a eu aussi une transmission nosocomiale connexe au Royaume-Uni chez un aide-soignant [13].
La possibilité d’une transmission zoonotique du MPXV a été démontrée de façon spectaculaire en 2003 dans les Etats du Midwest américain [14] lors de l’importation de cricétomes des savanes (Cricetomys gambianus), communément appelés rats de Gambie. Importés du Ghana, ces rats africains d’origine sauvage apparemment sains étaient vendus comme nouveaux animaux de compagnie (NAC) dans des animaleries où ils ont pu contaminer des chiens de prairie (Cynomys ludovicianus), autres rongeurs NAC autochtones de la famille des Sciuridae, qui furent les vecteurs secondaires d’une contamination humaine avec 71 cas, dont plusieurs enfants. Ce fut la seule épidémie importante de variole simienne d’origine zoonotique observée dans un pays non africain.
Rappelons que des rats NAC importés de Hongrie ont causé en Europe en 2009 une autre épidémie, celle-ci due au cowpox, un orthopoxvirus moins pathogène [15].
Du 7 mai 2022 au 30 juin 2022, le nombre des cas humains hors Afrique n’a pas cessé d’augmenter puisque l’on recense 5311 cas dans cinquante-trois pays non africains. En France, 498 cas ont été rapportés, dont une femme et un enfant.
Aspects cliniques
Le tableau clinique de la variole simienne se distingue de celui de la variole humaine par une adénopathie, absente dans la variole. L’éruption cutanée concerne le visage, les mains et différentes parties du corps : papules se transformant en vésicules puis en pustules, qui évoluent vers la cicatrisation avec formation de croûtes et des cicatrices cutanées moins graves que dans la variole humaine.
La variole simienne présente aussi des similitudes avec la varicelle, cette dernière étant toutefois plus contagieuse et causée par un herpèsvirus.
La maladie est généralement bénigne et il n’y a pas eu de mortalité en dehors d’un cas au Nigéria.
Transmissions
Les voies d'exposition potentielles comprennent l'interaction avec des animaux sauvages et la proximité d'individus malades, ainsi que le contact avec l’environnement contaminé.
Transmission zoonotique
Dès 1970, la variole du singe a été considérée comme une maladie virale émergente tout d’abord en RDC où les infections primaires humaines résultent d'un contact avec un animal sylvestre infecté, bien que l’espèce hôte réservoir soit actuellement inconnue.
Il peut s’agir d’un contact avec un animal sauvage devenu familier (rats de Gambie) ou chassé pour être consommé (viande de brousse).
Le seul MPXV isolé d'un mammifère sauvage a été obtenu à partir d'un écureuil à cordes moribond (Funisciurus anerythrus) collecté lors d'une enquête sur une épidémie au RDC [18, 19]. Les animaux les plus fréquemment suspects sont le Cricetomys gambianus (rat de Gambie), le Cercopithecus ascanius (singe à queue rousse) et les écureuils africains, en particulier le genre Funisciurus et Heliosciurus rufobrachium (écureuil soleil à pattes rouges) [16, 20].
Transmission interhumaine
La transmission interhumaine peut être directe, en particulier par le contact cutané (peau à peau), les fluides corporels et les voies respiratoires (gouttelettes). En raison de la grande résistance du virus, tout matériel (vaisselle...) ou linge ayant été en contact avec les croûtes d’un malade est contaminant.
Transmission des cas hors Afrique
Pour les cas inhabituels observés actuellement hors Afrique, les rencontres sexuelles ont joué clairement un rôle dans la transmission [21]. La maladie a surtout été observée chez des homosexuels masculins qui avaient participé à des réunions de leur communauté (fête en Belgique, sauna en Espagne, voyages...).
Moyens de lutte
Il importe évidemment d’éviter tout contact avec la personne atteinte et avec tout ce qu’elle a pu toucher et contaminer.
Les mesures d’éradication ne pourront pas être aussi efficaces avec la variole simienne qu’avec la variole humaine du fait d’un réservoir viral dans plusieurs populations d’animaux sauvages en Afrique.
Conclusion
Agent pathogène réémergent, le virus de la variole du singe semble ne plus se circonscrire aux régions endémiques africaines et le risque est de le voir coloniser mondialement la niche écologique laissée vacante par la variole.
La découverte soudaine de cette maladie hors de l’Afrique chez plus de 5000 personnes depuis le 7 mai 2022, sous une forme très particulière dans près de cinquante pays qui semblaient indemnes, avec un mode de transmission particulier, doit nous amener à une certaine prudence sur l’interprétation de cette émergence.
Cette maladie s'est-elle propagée pendant un certain temps sans être détectée du fait de son évolution bénigne ? Le 25 juin 2022, l’OMS n’envisage pas de déclarer la variole du singe en tant qu’urgence sanitaire mondiale. Mais plus ce virus se propagera longtemps et loin, plus il risque d’être endémique dans de nouvelles régions. Le problème sera de savoir si le MPXV s’établira plus dans un réservoir animal que dans la population humaine s’il continue à se propager.
L’important est de savoir diagnostiquer la variole du singe, d’avertir le public sur le risque de contagiosité de ce virus très résistant dans le milieu extérieur, de la possibilité de zoonose, de surveiller les cas contacts et de prévenir les contaminations des animaux sensibles (NAC exotiques). On peut espérer que la mise en place de ces mesures de biosécurité, associées ou non à une vaccination, limitera toute progression du MPXV en dehors de l’Afrique.
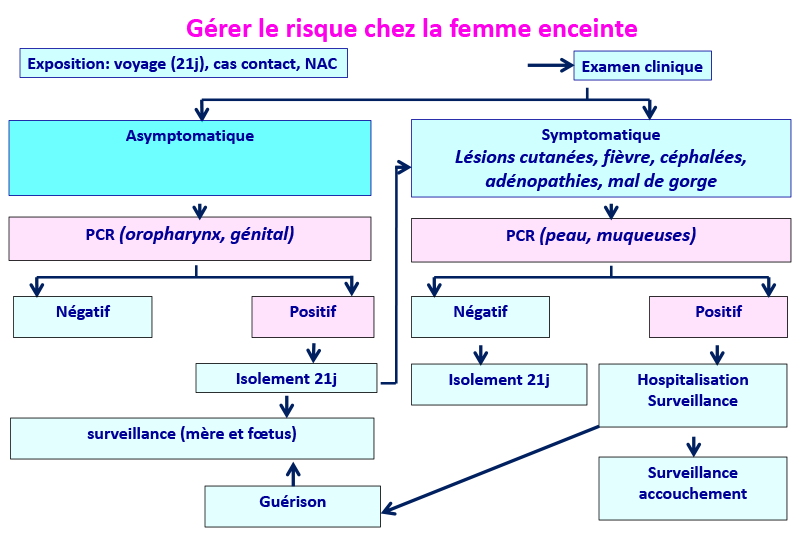
Ce schéma peut s’appliquer aussi à tous les cas d’infection par le MPXV.
(Modifié de www.thelancet.com. Publi. online June 21, 2022)
[1] Magnus P von, Andersen EK, Petersen KB, Birch-Andersen A. A pox-like disease in cynomolgus monkeys. Acta Pathologica Microbiologica Scandinavica. 1959;46(2):156‑76.
[2] Chastel C. Le monkeypox s’épidémise et franchit l’Atlantique (2003). In: Virus émergents. Vers de nouvelles pandémies ?. Vuibert. Adapt. Paris; 2006. 165‑76.
[3] Milhaud C, Klein M, Virat J. Analyse dʼun cas de variole du singe (monkeypox) chez le chimpanzé (Pan troglogdytes). Exp Anim. 1969;2:121‑35.
[4] Bunge EM, Hoet B, Chen L, Lienert F, Weidenthaler H, Baer LR, et al. The changing epidemiology of human monkeypox – A potential threat? A systematic review. Gromowski G, éditeur. PLoS Negl Trop Dis. 11 fév. 2022;16(2):e0010141.
[5] Gomes JP, Isidro J, Borges V, Pinto M, Sobral D, Santos J, et al. Multi-country outbreak of monkeypox virus: phylogenomic characterization and signs of microevolution [Internet]. In Review; 2022 mai [cité 26 juin 2022]. Disponible sur https://www.researchsquare.com/article/rs-1700947/v1
[6] Ladnyj ID, Ziegler P, Kima E. A human infection caused by monkeypox virus in Basankusu Territory, Democratic Republic of the Congo. Bull Org Mond Santé. 1972;46:593‑7.
[7] Durski KN, McCollum AM, Nakazawa Y, Petersen BW, Reynolds MG, Briand S, et al. Emergence of Monkeypox — West and Central Africa, 1970–2017. MMWR Morb Mortal Wkly Rep. 16 mars 2018;67(10):306‑10.
[8] World Health Organization. Disease Outbreak News; Multi-country monkeypox outbreak in non-endemic countries [Internet]. 2022 [cité 22 mai 2022]. Disponible sur https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON385
[9] Brown K, Leggat P. Human Monkeypox: Current State of Knowledge and Implications for the Future. Tropical Med. 20 déc 2016;1(1):8.
[10] Vaughan A, Aarons E, Astbury J, et al. Two cases of monkeypox imported to the United Kingdom, September 2018. Euro Surveill. 2018;pii=1800509.
[11] Erez N, Achdout H, Milrot E, Schwartz Y, Wiener-Well Y, Paran N, et al. Diagnosis of Imported Monkeypox, Israel, 2018. Emerg Infect Dis. mai 2019;25(5):980‑3.
[12] Ng OT, Lee V, Marimuthu K, Vasoo S, Chan G, Lin RTP, et al. A case of imported Monkeypox in Singapore. The Lancet Infectious Diseases. nov 2019;19(11):1166.
[13] Vaughan A, Aarons E, Astbury J, Brooks T, Chand M, Flegg P, et al. Human-to-Human Transmission of Monkeypox Virus, United Kingdom, October 2018. Emerg Infect Dis. avr 2020;26(4):782‑5.
[14] Centers for Disease Control and Prevention (CDC). Update: multistate outbreak of monkeypox – Illinois, Indiana, Kansas, Missouri, Ohio, and Wisconsin, 2003. MMWR Morb Mortal Wkly Rep. 11 juill 2003;52(27):642‑6.
[15] Campe H, Zimmermann P, Glos K, Bayer M, Bergemann H, Dreweck C, et al. Cowpox Virus Transmission from Pet Rats to Humans, Germany. Emerg Infect Dis. mai 2009;15(5):777‑80.
[16] Walker M. Monkeypox Virus Hosts and Transmission Routes: A Systematic Review of a Zoonotic Pathogen. Biological Sciences Undergraduate Honors Theses [Internet]. 1 mai 2022; Disponible sur https://scholarworks.uark.edu/biscuht/69
[17] Alakunle E, Moens U, Nchinda G, Okeke MI. Monkeypox Virus in Nigeria: Infection Biology, Epidemiology, and Evolution. Viruses. 5 nov 2020;12(11):1257.
[18] Reynolds MG, Carroll DS, Olson VA, et al. C. A Silent Enzootic of an Orthopoxvirus in Ghana, West Africa: Evidence for Multi-Species Involvement in the Absence of Widespread Human Disease. The American Journal of Tropical Medicine and Hygiene. 1 avr 2010;82(4):746‑54.
[19] Jezek Z, Grab B, Szczeniowski MV, Paluku KM, Mutombo M. Human monkeypox: secondary attack rates. Bull World Health Organ. 1988;66(4):465‑70.
[20] Monroe BP, Doty JB, Moses C, Ibata S, Reynolds M, Carroll D. Collection and Utilization of Animal Carcasses Associated with zoonotic Disease in Tshuapa District, the Democratic Republic of the Congo, 2012. Journal of Wildlife Diseases. juill 2015;51(3):734‑8.
[21] Endo A, Murayama H, Abbott S, Ratnayake R, Pearson CAB, Edmunds WJ, et al. Heavy-tailed sexual contact networks and the epidemiology of monkeypox outbreak in non-endemic regions, May 2022 [Internet]. Epidemiology; 2022 juin [cité 26 juin 2022]. Disponible sur http://medrxiv.org/lookup/doi/10.1101/2022.06.13.22276353
[22] PRO/AH/EDR> Monkeypox update (01) [Internet]. ProMED-mail. 2022 [cité 21 mai 2022]. Disponible sur https://promedmail.org/
[23] Promed. Monkeypox: Health ministry confirms only one person contracted virus [Internet]. Promed; 2022 [cité 22 mai 2022]. Disponible sur https://www.jpost.com/breaking-news/article-707339
Christian Marchal
Astronome

«Suspendez votre cours :
«Laissez-nous savourer les rapides délices
«Des plus beaux de nos jours !»
En quelques lignes tout est dit, mais les regrets du poète ne trompent personne, pas même lui. Le temps se rit de nous et nous échappe perpétuellement ; ses rapports avec l’humanité sont les plus complexes et les plus élaborés qui soient. L’invention de l’heure est la première tentative pour dompter le temps, mais ce n’est pas un absolu, c’est une convention nécessaire à l’organisation de la vie en société. Les multiples modifications historiques de la définition de l’heure prouvent son caractère artificiel...
Heure d’été, heure d’hiver, alternance ou pas ? La question est largement débattue. L’heure d’été gardée toute l’année aurait la préférence, dit-on, mais les inconvénients de l’heure d’été en hiver sont beaucoup plus graves que ceux de l’heure d’hiver en été : levez-vous par la nuit noire et allez au travail à l’heure la plus froide sur des routes verglacées... Le plus étonnant est que cette mésaventure nous est déjà arrivée dans les années qui ont suivi la Libération, mais nous avons oublié...
Le temps solaire «vrai» et le temps solaire moyen
Plantez un bâton verticalement un jour ensoleillé, son ombre tournera peu à peu. Vous pouvez marquer sur le sol la position de l’ombre pour les heures successives et vous constaterez le lendemain que l’ombre revient presque exactement aux mêmes places à chaque heure.
Le bâton vertical et son ombre sont la première esquisse des cadrans solaires mais pour obtenir une excellente régularité, il convient d’utiliser une tige parallèle à l’axe de rotation de la Terre (donc inclinée en direction de l’étoile polaire) ; cet élément essentiel a été découvert, de manière le plus souvent pragmatique, par toutes les civilisations de l’Antiquité. La tradition est de mettre «midi» (en latin le milieu du jour) dans le plan vertical de la tige : il est midi quand le soleil est au plus haut et pour éviter toute confusion, les astronomes appellent cet instant «midi vrai» ; le temps correspondant est le «temps vrai».
La construction d’horloges précises a révélé que le «temps vrai» n’est pas rigoureusement régulier, car l’orbite de la Terre n’est pas tout à fait circulaire et son axe est incliné ; le temps moyen correspondant s’appelle «temps solaire moyen». La différence temps moyen moins temps vrai est fonction des saisons mais reste toujours petite ; elle est maximale le 4 février (14mn 18s) et minimale le 3 novembre (retard de 16mn 24s).
A partir de 1816, toutes les horloges publiques de France sont réglées sur le temps solaire moyen local et chacun «voit midi à sa porte», mais le développement des chemins de fer fait prendre conscience que tous ces temps sont différents : Strasbourg est en avance de 21mn 40s sur Paris et Brest en retard de 27mn 20s. La confusion générée par cette situation est telle que les différents réseaux de chemin de fer se mettent à utiliser partout l’heure de Paris et, pendant près de vingt ans, il y aura dans chaque province «l’heure de la ville et l’heure de la gare». Enfin, en mars 1891, toute la France métropolitaine se met à l’heure de la capitale. La plupart des pays font de même et il vous faut changer d’heure chaque fois que vous traversez une frontière : il faut retarder votre montre de 9mn 21s si vous traversez la Manche. La clarté y a gagné, mais c’est encore bien compliqué et le vrai progrès sera l’adoption du système des fuseaux horaires. Les Français rechigneront longtemps à adopter ce système basé sur le méridien de Greenwich et sur le «temps moyen de Greenwich» appelé GMT ou TU (temps universel) et ce n’est qu’en mars 1911 que l’heure légale en France sera définie comme étant «l’heure du temps moyen de Paris, diminuée de 9mn 21s». La perfide Albion n’est pas mentionnée, l’honneur national est sauf !
L’heure d’été
«De vent, de froidure et de pluie,
«Et s’est vêtu de broderie,
«De soleil luisant, clair et beau.»
Les Scandinaves en conviennent volontiers : «Nous vivons au nord, mais nous portons l’été dans nos cœurs !». Tous les peuples qui vivent loin de l’équateur sont conditionnés par l’alternance des saisons : la belle saison chaude aux longues journées et la saison froide aux nuits interminables. Certes les différences ne sont pas partout aussi marquées qu’en Scandinavie, mais déjà à 30° de latitude (Le Caire, la Floride), une journée moyenne des quatre mois de mai à août est trois heures plus longue qu’une journée moyenne des quatre mois de novembre à février ; la différence atteint quatre heures à Alger et cinq heures à Rome, elle dépasse six heures à Paris... Les deux saisons opposées sont vraiment différentes !
L’idée d’avancer les horloges en été, à la fois pour économiser la lumière et nous rapprocher de nos rythmes naturels – nos ancêtres se levaient comme le soleil – fut exprimée par William Willet, qui mourut en 1915, un an à peine avant qu'elle soit appliquée.
1914. Une fois encore, la vieille Europe est ensanglantée. Mais cette fois-ci, les peuples entiers sont au combat, on ne se contente plus d’armées de 30 000 hommes comme au XVIIIe siècle. Les armées mobilisent des millions d’hommes et dévorent en quelques mois des ressources que l’on avait mis des décennies à accumuler. Dans ces conditions, André Honorat, député des Basses-Alpes, n’a pas trop de peine à convaincre ses collègues de l’intérêt de l’idée de William Willet et la France adopte l’heure d’été le 14 juin 1916, un mois à peine après la Grande-Bretagne.
L’alternance TU en hiver et TU + 1h en été va être jugée très intéressante et conservée une fois la paix revenue. Les changements auront toujours lieu dans la nuit de samedi à dimanche, en général à la fin de mars et à la fin de septembre.
1940. L’invasion, l’occupation. La situation va être chaotique, elle dépendra des zones d’occupation et des flux et reflux des armées.
1945. La guerre est finie. Va-t-on reprendre l’alternance de l’avant-guerre ? La question soulève des discussions passionnées. Finalement l’alternance est abandonnée en 1946 et l’on garde toute l’année l’heure d’été de l’avant-guerre (TU + 1h) au motif que cette heure permettra les économies d’énergie associées à l’heure d’été, tout en évitant le changement d’heure tous les six mois. Mais bien vite cela se révèle une illusion, c’est l’alternance et les horaires qui y sont adaptés qui entraîne les économies, d’autre part l’heure d’été utilisée en hiver a de graves inconvénients : levez-vous par la nuit noire des petits matins de décembre ou janvier et allez au travail à l’heure la plus froide sur des routes verglacées... Très vite, tous ceux qui le peuvent retardent leurs horaires et au bout de quelques années, pratiquement tous les horaires ont été retardés d’une heure. La France métropolitaine gardera cette heure, TU + 1h (modernisée en UTC + 1h) jusqu’en 1975.
1973. Le premier choc pétrolier. Le prix du baril passe de 4 à 16 dollars en quelques semaines. Chacun cherche à faire des économies et l’on finit par se rappeler que l’alternance heure d’été heure d’hiver est un bon moyen d’en faire pour pratiquement aucun investissement. Pour la France, l’économie correspond à la consommation énergétique d’une ville de 800 000 habitants. Oui mais quelle heure choisir ? Va-t-on reprendre les rythmes d’avant-guerre alors que la plupart des horaires ont été retardés d’une heure ? La solution de sagesse sera de ne pas modifier les horaires et de prendre UTC + 1h en hiver et UTC + 2h en été, ce qui a d’autre part l’avantage d’unifier dans ce domaine le continent européen : une heure identique de l’Espagne à la Pologne et de la Norvège à l’Italie et la Yougoslavie.
Cette situation laborieusement obtenue sera-t-elle pérenne ? Pas si sûr, plusieurs professions soulignent les inconvénients des changements d’heure : ainsi les vaches laitières habituées à la traite de 19h supportent très mal de devoir attendre une heure de plus et les éleveurs sont contraints d’étaler le changement...
Projet d’heure naturelle
Soulignons d'entrée que le projet qui va suivre n'est pas actuellement applicable ; il le sera cependant le jour où les horloges seront numériques et réglées automatiquement comme celles des ordinateurs. Ainsi la sophistication numérique extrême rejoindrait-elle la nature. Compte-tenu des diverses contraintes, simplicité, économie, proximité des rythmes naturels (nos ancêtres se levaient bien plus tôt en été qu’en hiver), absence de décalages brutaux, on peut envisager le système suivant que l’on peut appeler «heure naturelle» :
- A. Du 31 octobre au 31 mars, on utilise l’heure d’hiver habituelle.
- B. En avril, on passe en douceur à l’heure d’été en ne mettant que 58mn chaque matin, entre 2h et 3h.
- C. De mai à septembre, on utilise l’heure d’été habituelle.
- D. Enfin, on repasse à l’heure d’hiver en mettant 62mn entre 2h et 3h chaque matin, du 1er au 30 octobre. On pourra ainsi se donner rendez-vous à 2h61...
Le caractère naturel de ce projet peut être souligné par le fait suivant : aux latitudes moyennes de l’hémisphère nord, entre 45° et 50°, le soleil se lève pratiquement à la même heure tout au long du mois d’avril (à Paris entre 6h33 et 6h35 avec les heures d’été et d’hiver de ce début de XXIe siècle. Pour octobre, la zone d’équilibre est un peu plus au nord.
Conclusion
L’homme est mortel, le temps finit toujours par gagner. L’heure et le calendrier sont des moyens inventés par l’humanité pour tenter de dompter et de s’approprier cet insaisissable que nous appelons le temps et les complications extrêmes de ces tentatives démontrent à l’envie leur importance. Pensez aux avatars du 29 février, pensez que tant de peuples se sont construit un calendrier et l’ont peu à peu affiné au fil des siècles... Le mètre et le kilomètre sont la simplicité même à côté du jour, de la semaine, du mois, de l’année, de l’heure, de la minute, de la seconde !
Dans ces conditions, il est logique que nos systèmes de mesure du temps aient profondément varié au cours des siècles, tiraillés entre la régularité absolue – mais contraire à nos rythmes naturels – et une adaptation brutale – une heure de plus ou de moins d’un coup –, coincés entre les habitudes locales et les nécessités de liaisons globales... On peut espérer que l’heure naturelle permettra de concilier tous ces extrêmes.
GMT : temps moyen de Greenwich (Greenwich Mean Time)
TU : temps universel, autre nom du GMT
UTC : temps universel coordonné : toujours très voisin du TU (écart < 0,9s) mais défini à l’aide du temps atomique international, temps bien plus régulier que la rotation de la Terre.
Claude Monneret* et Caroline Victorri-Vigneau**
*Directeur de recherche honoraire au CNRS, ancien président de l’Académie nationale de pharmacie
** Professeure des Universités – praticienne hospitalière au CHU de Nantes, Inserm, UMR 1246 «SPHERE» (MethodS in Patients-centered outcomes and HEalth REsearch)
Article issu de la conférence du 8 mars 2022 à l'Institut Curie (Partenariat AFAS / Chercheurs Toujours), paru dans la Lettre de Chercheurs Toujours, N° 38, mai 2022.

Le protoxyde d’azote est un gaz incolore, utilisé comme gaz propulseur (E942) notamment dans les cartouches destinées aux siphons alimentaires. Mais c’est aussi un gaz à usage médical, utilisé principalement pour ses propriétés analgésiques. Il est utilisé comme anesthésique de courte durée, en unité de chirurgie et d'urgence et surtout en pédiatrie, associé à l’oxygène (mélange 50/50) sous le nom de MEOPA. De façon beaucoup plus récente, il a maintenant des indications en cardiologie pour ses propriétés analgésique et anxiolytiques.
De l’attraction dans les foires à l’usage médical : la sérendipité
Découvert en 1772 par le chimiste anglais Joseph Priestley, ses propriétés euphorisantes ne seront découvertes que vingt-cinq ans plus tard par un autre chimiste anglais, Humphry Davy. Celui-ci, vers 1800, l’étudie de manière détaillée, tant au niveau de ses propriétés physiques et chimiques que de ses effets lorsqu’il est inhalé. Davy découvre ainsi les propriétés euphorisantes du gaz, mais également ses vertus anesthésiantes, qu’il résume ainsi : «As nitrous oxide in its extensive operation appears capable of destroying physical pain, it may probably be used with advantage during surgical operations in which no great effusion of blood takes place»
Les conclusions de Davy concernant les effets analgésiques de l’oxyde nitreux furent laissées de côté. Il n’y eut que Stodart (1802) et Barton (1808) qui constatèrent aussi les effets analgésiques du protoxyde d’azote mais sans proposer son usage dans un cadre chirurgical.
Rapidement, l’intérêt de ce gaz s’échappe des seuls cercles scientifiques et littéraires pour gagner les salons et les foires britanniques, où il devient le divertissement à la mode. L'engouement est tel que la pratique menace bientôt de se muer en problème de santé publique et doit être réglementée. Si la «gazomanie» touche l’Angleterre de plein fouet, elle ne semble atteindre la France que dans une moindre mesure. Tel est l’engouement que le cinéma s’en emparera, dont Charlie Chaplin.
Thomas McLean, Public domain, via Wikimedia Commons

Le protoxyde d'azote est alors utilisé en cette fin du XVIIIe siècle comme «gaz hilarant» dans les foires. Alors qu’il assiste à une démonstration, un des spectateurs, invité à monter sur l’estrade pour expérimenter le gaz, fait une chute en redescendant de l’estrade, se blessant profondément. A la grande surprise, il n’éprouve aucune douleur. Le dentiste Horace Wells, qui assiste à la démonstration, comprend aussitôt que le protoxyde d’azote est la raison de cette absence de réaction à la douleur. Pour vérifier cette hypothèse, il se fait extraire, dès le lendemain, une molaire en train de se gâter tandis qu’on lui administre le gaz. Le résultat est probant. Il renouvelle cette expérience avec succès auprès de quinze de ses patients au cours des mois suivants.
A Boston, Wells rencontre le Pr. Warren, chirurgien du Massachusetts General Hospital et tout un auditoire pour convaincre ses confrères dentistes mais également les médecins présents, des vertus anesthésiques du protoxyde d’azote. La séance se déroule le 20 janvier 1845 et Wells demande à son auditoire si quelqu'un a besoin de se faire extraire une dent, Wells commence sa démonstration avec l’assistance de son collègue Morton, mais suite à une mauvaise administration du gaz, l'étudiant volontaire se plaint d'avoir très mal.
Suicide d’Horace Wells
Cette expérience désastreuse marqua un sérieux coup d'arrêt dans l'utilisation du protoxyde d'azote en anesthésie et discrédita Wells aux yeux de la communauté médicale, allant le qualifier même de charlatan. Rentré dépressif à Hartford, Horace Wells ne rouvrit pas son cabinet, vivant chichement du produit de ses conférences sur l’ornithologie. Par la suite, devenu addict au chloroforme, Horace Wells, en plein délire, attaque le 21 janvier 1848 deux prostituées sur lesquelles il jette de l’acide sulfurique. Arrêté, il met fin à ses jours le 24 janvier 1848 en se tranchant l’artère fémorale, laissant une lettre pour expliquer son geste.
Protoxyde d‘azote, éther ou chloroforme ?
Parallèlement aux essais de Wells, d’autres vont préconiser l’utilisation de l’éther et c’est ainsi qu’il faudra attendre les recherches et démonstrations réussies avec l'éther de William Thomas Green Morton, un élève de Wells en collaboration avec le Dr John Warren (octobre et novembre 1846 à Boston) pour que l'anesthésie soit enfin utilisée par les chirurgiens. En France, c’est le chirurgien Antoine-Joseph de Lamballe qui opérera à l’hôpital Saint-Louis le premier patient sous anesthésie à l’éther, le 22 décembre 1846. Toutefois, plus simple à manipuler, le chloroforme sera préféré durant la guerre de 1871.
En 1864, le dentiste John W. Crane introduit le protoxyde d’azote en France. Deux ans plus tard, Apolloni-Pierre Préterre, un chirurgien-dentiste apporte quelques perfectionnements importants au gazomètre et au masque d’inhalation. Crane et Préterre seront donc les premiers utilisateurs du N2O tout en rappelant que les anesthésies à l’éther ou au chloroforme sont responsables de mortalités et qu’un nouvel anesthésique est à rechercher, ajoutant... «nous l’avons trouvé, c’est le protoxyde d’azote».
Tout fut alors question d’antériorité concernant le «découvreur de l’anesthésie chirurgicale» entre Wells et Morton. Bien plus tard, la France honora Wells en reconnaissant son antériorité dans la découverte de l’anesthésie chirurgicale et une statue fut érigée au 1 square des Etats-Unis à Paris. Sur le socle, on peut lire : «Au dentiste Horace Wells, novateur de l’anesthésie chirurgicale».
Luca Borghi, CC BY-SA 4.0, via Wikimedia Commons

Euphorie, analgésie : une pharmacologie complexe
Au niveau du cerveau, le protoxyde d’azote agit sur de nombreux circuits impliqués dans la nociception, l’anxiolyse et entraîne une libération accrue de dopamine, le neurotransmetteur du circuit de récompense, ce qui explique ses multiples effets, notamment anxiolytiques, analgésiques et euphorisants à l’origine de son appellation de «gaz hilarant». Le protoxyde d’azote est utilisé comme anesthésique de courte durée, en unité de chirurgie et d'urgence et, surtout en pédiatrie, associé à l’oxygène (mélange 50/50) sous le nom de MEOPA. De façon beaucoup plus récente, il a maintenant des indications en cardiologie pour ses propriétés analgésique et anxiolytique. Toutefois, selon le dictionnaire de l’Académie nationale de médecine, le rire inextinguible et l'amnésie consécutifs à l'inhalation de N2O sont dûs à l'hypoxie aigüe transitoire produite par l'administration de N2O pur et non à l'anesthésie.
Détournement d’usages et conséquences physiologiques
L’inhalation du gaz contenu dans les cartouches est une pratique en vogue chez les jeunes. Au début, le protoxyde d’azote était cantonné à l’espace festif, mais depuis 2018, la consommation de protoxyde d’azote a explosé. C’est ainsi qu’aujourd’hui on observe des consommations répétées, voire quotidiennes, de très grande quantités qui sortent totalement du milieu festif. Les effets recherchés par les usagers de ce gaz sont principalement l’euphorie, mais aussi la distorsion des perceptions auditives ou visuelles, les sensations de dissociation, désinhibition, «flottement» et également des recherches de sensation de bien-être. La durée de ces effets est très courte. Elle ne dépasse pas quelques minutes. Ce qui peut conduire à des prises répétées du produit, d’où une réelle addiction avec les signes de dépendance conduisant à une augmentation des doses pour obtenir l’effet recherché et a priori, en cas d’arrêt de consommation, un sevrage.
Au passage, Caroline Vigneau souligne que la pharmacodépendance est un problème de santé publique, car les Français se situent parmi les champions des consommateurs de psychotropes, de benzodiazépines et de cannabis, De là, le rôle essentiel des centres d’addictovigilance qui est d’évaluer le risque pour le consommateur et comment le prévenir en santé publique. Ces centres scrutent également l’émergence de nouvelles tendances en la matière. Celui des Pays de la Loire est plus spécifiquement responsable de la surveillance du protoxyde d’azote qui, paradoxalement, fait l’objet d’une surveillance étroite sous sa forme pure (classé en liste 1 des substances vénéneuse). Utilisé dans les blocs opératoires sous forme de MEOPA, il permet la prise en charge de patients. En odontologie pédiatrique, il permet de soigner les patients handicapés ou anxieux. Il répond à la classification des stupéfiants alors qu’en tant qu’additif alimentaire E942, il est librement distribué. Ce paradoxe vient du fait que la version médicamenteuse est gérée par l’Agence du médicament, qui assume son suivi et sa sécurité, alors que le E942 dépend de l’Anses (Agence nationale de sécurité de l’alimentation, de l’environnement et du travail).
Des risques sur la santé
Cette évolution des consommations a été accompagnée d’une augmentation du nombre de signalements d’effets sanitaires graves. Le premier risque est l’asphyxie par manque d’oxygène. Sont aussi décrits notamment des brûlures par le froid du gaz lorsqu’il est expulsé de la cartouche, des pertes de connaissance, des vertiges, un risque de chute important, de désorientation et d’accidentologie. Tous ces effets peuvent apparaître juste après l’inhalation.
En cas d’utilisation répétée ou de prise de fortes doses, des atteintes neurologiques peuvent survenir, dont des cas graves avec atteintes du système nerveux central et de la moelle épinière pouvant entraîner des conséquences irréversibles. L’arrêt des consommations et un diagnostic immédiat, avec une prise en charge thérapeutique en neurologie, peuvent limiter les risques. Tout signe neurologique après une inhalation doit conduire à une consultation la plus rapide possible. D’autres conséquences cliniques, notamment psychiatriques et cardiaques, ont été rapportées à la suite d'une consommation de protoxyde d’azote.
Enfin, il existe des cas de troubles de l’usage et de dépendance. Les fréquences et les quantités consommées demeurent variables mais aujourd’hui certains sujets consomment jusqu’à plusieurs centaines de cartouches par jour.
Une surveillance d’addictovigilance rapprochée
Les centres d’addictovigilance, au sein desquels travaillent des pharmacologues médicaux, dont les missions sont coordonnées par l’Agence nationale de sécurité du médicament et des produits de santé (ANSM) ont pour mission de recueillir les cas et d’évaluer le potentiel d’abus et de dépendance des substances, afin de prévenir le risque en santé publique. Ils alertent depuis plusieurs années sur détournement du protoxyde d’azote.
La loi n°2021-695 du 1er juin 2021 tendant à prévenir les usages dangereux du protoxyde d'azote établit un cadre protecteur en prévoyant «l’interdiction de vendre ou d'offrir du protoxyde d'azote aux mineurs, quel que soit le conditionnement, dans tous les commerces, les lieux publics et sur Internet». Il reste toutefois important de sensibiliser tous les professionnels de santé concernés par ce phénomène ainsi que les associations d’usagers afin d’optimiser l’information, la prévention, le repérage et la prise en charge des sujets.
Devançant la loi, un certain nombre de municipalités ont pris des arrêtés, comme celui du 12 octobre 2020 à Limeil-Brévannes, pour interdire la vente du protoxyde d’azote aux mineurs et sa détention ou consommation sur l’espace public.
Jeanne Brugère-Picoux
Professeur honoraire de pathologie médicale du bétail et des animaux de basse-cour (Ecole nationale vétérinaire d’Alfort), membre de l’Académie nationale de médecine, présidente honoraire de l’Académie vétérinaire de France
Depuis le 6 mai 2022, nous assistons à une émergence inhabituelle de cas humains sporadiques de la variole du singe dans plusieurs pays européens, en Amérique du Nord et en Australie alors qu’il s’agissait d’une maladie localisée à l’Afrique centrale et occidentale. La particularité de ces cas est que, en dehors du premier malade au Royaume-Uni de retour du Nigéria, il s’agit de cas autochtones n’ayant pas voyagé en Afrique. Alors que l’on reconnaît une origine zoonotique dans la plupart des cas africains, ces cas émergents sont liés à des contaminations interhumaines, souvent observées chez des hommes homosexuels ou bisexuels présentant des lésions cutanées génitales et au niveau du visage. C’est pourquoi une transmission sexuelle peut être suspectée. La transmission interhumaine est possible par le contact avec les fluides corporels, les lésions cutanées (dont les croûtes), l'environnement ou les objets contaminés par le malade. Il peut aussi s’agir d’une contamination d’origine nosocomiale.
Le nombre de ces cas autochtones émergents détectés depuis deux semaines a dépassé largement l’ensemble des cas importés par des voyageurs revenant du continent africain depuis 1970 puisque près d’une centaine de cas sont répertoriés au 22 mai 2022 (cf. encart). De nombreuses autorités sanitaires et l’OMS craignent une augmentation des cas rapportés pendant la période estivale (où les gens se rassemblent pour des festivals et des fêtes). en raison de la propagation silencieuse de ce virus variolique, apparu simultanément dans des régions géographiquement éloignées, chez des personnes sans lien apparent.
Seule une enquête épidémiologique complète nous permettra d’évaluer le risque lié à ce virus émergent hors de sa région géographique habituelle. Ce nouveau comportement viral peut être inquiétant, notamment si un risque zoonotique, non démontré à ce jour dans ces cas récents, ne peut être formellement exclu chez une espèce animale non africaine.
Au 23 mai 2022, la variole du singe a été identifiée sous une forme bénigne sans mortalité dans 15 pays avec près de 100 cas confirmés, et d’autres cas sont suspectés [1, 2, 3].
Europe :
- Espagne : 30 cas ; les autorités ont fermé un sauna dans la région de Madrid qui semble lié à plusieurs contaminations chez des hommes homosexuels ;
- France : un cas annoncé le 20 mai 2022 (homme de la région Ile-de-France n’ayant pas voyagé récemment) ;
- Belgique : 2 cas confirmés ayant participé à une même fête ;
- Allemagne : 1 cas rapporté ;
- Italie : 3 cas annoncés le 20 mai 2022 ;
- Pays-Bas : 1 cas ;
- Suède : 1 cas annoncé le 19 mai 2022 dans la région de Stockholm ;
- Portugal : 14 cas et 20 autres sous surveillance ;
- Royaume-Uni : d’abord 9 cas signalés depuis le 6 mai 2022 ; les autorités sanitaires anglaises ont annoncé 11 cas supplémentaires le 20 mai 2022 et craignent l’apparition de nouveaux cas et signalent que pour certains, il s’agit de contaminations chez des hommes homosexuels ou bisexuels ;
- Suisse : 1 cas observé le 22 mai 2022 chez une personne ayant été contaminée à l’étranger ;
- Grèce : un cas annoncé le 22 mai 2022 chez un touriste anglais.
Moyen-Orient :
- Israël : 1 cas a été annoncé le 22 mai 2022 chez un homme de 30 ans ayant eu un contact avec un malade à l’étranger.
Amérique du Nord :
- Canada : 2 cas signalés et 17 autres cas suspects à Montréal ;
- Etats-Unis : 1 cas dans l'Etat du Massachusetts signalé chez un homme qui avait récemment voyagé au Canada.
Pacifique :
- Australie : 2 cas probables chez des voyageurs en provenance d’Europe, à Melbourne (30 ans) et à Sydney (40 ans).
Découverte du virus de la variole du singe en 1958 puis de la variole simienne chez l’Homme en 1970 en Afrique
Le virus de la variole du singe a été découvert en 1958 chez des singes de laboratoire à Copenhague mais ce n’est qu’en 1970 qu’un premier cas humain a été décrit en République démocratique du Congo (RDC) chez un enfant âgé de 9 mois qui n’avait pas eu de contact démontré avec des singes [4]. Il est possible que cette infection ait été masquée avant 1970 dans ce pays africain du fait d’une confusion possible avec la variole humaine qui était encore endémique en Afrique. L’éradication de la variole humaine dans le monde avait été annoncée en 1977, ce qui a permis l’arrêt de la vaccination contre cette maladie.
Cette variole simienne ou monkeypox est différente de la variole humaine dans son tableau clinique observé après 6 à 13 jours d’incubation : une phase initiale non spécifique avec hyperthermie, adénopathie (absente dans la variole), myalgie, suivie d’une phase d’éruption cutanée (papules se transformant en vésicules puis en pustules évoluant vers une cicatrisation avec la formation de croûtes) sur le visage, les mains et différentes parties du corps.
Les cicatrices cutanées pouvant être observées sont moins graves que dans la variole humaine. La variole simienne ressemble aussi beaucoup à la varicelle, qui est plus contagieuse.
Le taux de létalité varie selon les deux souches virales connues : la souche Congo (ou souche d’Afrique centrale), la plus virulente, peut provoquer une mortalité de 10,6% alors que la souche d’Afrique occidentale est moins pathogène avec un taux de mortalité estimé de 3,6% [5]. La bonne nouvelle est que la souche isolée dans les cas actuels préoccupants serait la souche d’Afrique occidentale, ce qui explique les symptômes relativement bénins observés. Mais la mauvaise nouvelle est que ces cas bénins ne sont peut-être que la partie émergée d’un iceberg, pouvant faire craindre une sous-estimation du nombre de personnes infectées.
Maladie zoonotique virale émergente
Dès 1970, la variole du singe a été considérée comme une maladie virale émergente avec plusieurs épisodes en Afrique où, contrairement à la variole humaine, la contamination n’est pas uniquement interhumaine mais elle peut reconnaître une origine zoonotique (animal familier ou viande de brousse) du fait d’un portage souvent asymptomatique du virus variolique par de nombreux rongeurs, plus souvent en cause dans la transmission du virus à l’Homme que les primates non humains. Les espèces les plus souvent incriminées sont des écureuils africains, en particulier le genre Funisciurus, et le rat géant de Gambie (Cricetomys gambianus) [6, 7]. Ce poxvirus peut aussi contaminer de nombreuses espèces animales autres que des rongeurs ou des singes (hérisson, porc, éléphant, opossum...) [7]. Mais l’animal réservoir principal et définitif du virus n’a jamais été identifié en Afrique avec précision et l’on a jamais observé de transmission de l’Homme à l’animal [7].
Cette possibilité de transmission zoonotique a d’ailleurs été démontrée de façon spectaculaire en 2003 lors de l’importation de rats de Gambie dans les Etats du Midwest américain. Ces rats africains, apparemment sains, vendus comme nouveaux animaux de compagnie (NAC) ont pu contaminer dans l’animalerie des chiens de prairie, autres rongeurs NAC autochtones et qui furent les vecteurs secondaires d’une contamination humaine avec plus de 70 cas, dont plusieurs enfants. Ce fut la seule importante épidémie de variole simienne observée dans un pays non africain. Rappelons qu’en Europe nous avons connu en 2009 une autre épidémie due à un orthopoxvirus moins pathogène (le cowpox) avec des rats importés de Hongrie en tant que NAC.
Epidémiologie de la variole simienne
Au cours des années suivant sa première identification en Afrique, la variole du singe, qui n’était plus masquée par la variole (ou la vaccination), a été régulièrement observée en Afrique centrale et occidentale.
Identifiée au Nigéria dès 1971, la maladie a été très sporadique entre 1971 et 1978 (10 cas rapportés) puis on a pu noter une augmentation progressive du nombre des malades avec plusieurs milliers de cas confirmés dans 15 pays différents, dont 11 dans des pays africains (avec un taux de létalité d'environ 3 à 6%) et de rares cas sporadiques importés au Royaume-Uni, aux Etats-Unis, en Israël et à Singapour [7]. Les augmentations les plus spectaculaires se sont produites en RDC où, par exemple, il y a eu 1238 cas de variole du singe (dont 57 décès) entre le 1er janvier et le 1er mai 2022 [2]. L’âge médian des malades est passé de 4 ans dans les années soixante-dix à 21 ans depuis 2010 [5]. Cette augmentation est due à plusieurs facteurs : déclin de l'immunité collective à la suite de l'arrêt de la vaccination contre la variole permettant l’augmentation des contaminations interhumaines, changement climatique et déforestation favorisant une urbanisation des zones où le virus est présent dans ses réservoirs animaux permettant une contamination de l’Homme par l’animal, consommation de viande de brousse.
Moyens de lutte
Il est évident que dans les cas de cette poxvirose, il importe d’éviter tout contact avec la personne atteinte ainsi que tout ce qu’elle a pu toucher.
Par comparaison avec la variole humaine, les mesures d’éradication ne pourront pas être aussi efficaces avec la variole simienne du fait d’un réservoir viral dans plusieurs populations d’animaux sauvages en Afrique.
La vaccination contre la variole permettait d’offrir une protection croisée contre les orthopoxvirus, dont le virus de la variole simienne qui est estimée à 85% [8]. Les cas humains identifiés de variole du singe étaient naïfs pour 90% car beaucoup sont nés après l’arrêt de la vaccination [8]. Lors de l’épidémie de 2003 aux Etats-Unis, le vaccin variolique avait été recommandé par le Center for Disease Control and Prevention (CDC) pour réduire la gravité des symptômes mais il n’a pas empêché la maladie [8]. Il existe maintenant des vaccins de troisième génération, plus atténués, pouvant être utilisés chez les sujets immunodéficients à risque en Afrique. Les autorités sanitaires du Royaume-Uni ont annoncé la possibilité d’une vaccination «en anneau» si le nombre de cas humains augmentait.
Conclusion
Le virus de la variole du singe est un agent pathogène réémergent, qui ne semble plus se limiter aux régions endémiques africaines et qui présente le risque mondial d’occuper la niche écologique laissée vacante par la variole.
Il faut espérer que :
- cette transmission restera interhumaine et limitée ;
- qu’il n’y aura pas de cas humains asymptomatiques permettant une persistance du virus et sa propagation, avec une augmentation des cas confirmés dans les pays déjà atteints ;
- que cette apparente plus grande contagiosité du virus hors d’Afrique n’est pas liée à un nouveau virus qui aurait muté en devenant plus contagieux ;
- qu’il n’y aura pas un relais avec un réservoir animal autochtone dans les pays touchés pour cet agent zoonotique, comme c’est le cas en Afrique.
[2] World Health Organization. Disease Outbreak News; Multi-country monkeypox outbreak in non-endemic countries [Internet]. 2022 [cité 22 mai 2022]. Disponible sur: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON385.
[3] Promed. Monkeypox: Health ministry confirms only one person contracted virus. Promed; 2022.
[4] Ladnyj ID, Ziegler P, Kima E. A human infection caused by monkeypox virus in Basankusu Territory, Democratic Republic of the Congo. BullOrg.mond Santé. 1972;46:593‑7.
[5] Bunge EM, Hoet B, Chen L, Lienert F, Weidenthaler H, Baer LR, et al. The changing epidemiology of human monkeypox—A potential threat? A systematic review. Gromowski G, éditeur. PLoS Negl Trop Dis 11 févr 2022;16(2):e0010141.
[6] Walker M. Monkeypox Virus Hosts and Transmission Routes: A Systematic Review of a Zoonotic Pathogen. Biological Sciences Undergraduate Honors Theses [Internet]. 1 mai 2022; Disponible sur: https://scholarworks.uark.edu/biscuht/69.
[7] Alakunle E, Moens U, Nchinda G, Okeke MI. Monkeypox Virus in Nigeria: Infection Biology, Epidemiology, and Evolution. Viruses. 5 nov 2020;12(11):1257.
[8] Brown K, Leggat P. Human Monkeypox: Current State of Knowledge and Implications for the Future. Tropical Med. 20 déc 2016;1(1):8.
Denis Monod-Broca
Ingénieur et architecte, secrétaire général de l'Afas

L’Afas a rendu compte du dernier rapport du GIEC sous la plume d’Alain Foucault, en août, mars et avril derniers lors de la sortie des différentes parties de ce sixième rapport.
Un mot mérite qu’on s’y arrête. Les scientifiques du GIEC, dans leurs recommandations - et c’est une nouveauté qui a été relevée - mentionnent la sobriété.
Ce simple mot de sobriété, dans sa sobriété même, toute descriptive, toute scientifique, en dit long. Ses implications sont multiples et considérables.
«Je suis sobre désormais», dit l’ex-alcoolique qui a su vaincre son assuétude. N’est-ce pas cet exemple-là que les rédacteurs du rapport nous invitent à suivre ? Pour limiter le réchauffement climatique et, aussi, pour préserver les ressources naturelles, y a-t-il une autre voie que celle qui consiste à vaincre notre addiction au «toujours plus» ?
Beaucoup de pays, beaucoup d’individus à la surface de la Terre ont à peine de quoi vivre, il ne s’agit pas de leur demander, à eux, d’être sobres, ils le sont, ils ne le sont que trop. Mais nous, nous pays riches, nous riches habitants des pays riches, nous avons trop et - comment en douter ? - nous devons nous désintoxiquer de ce trop. Quoi de plus difficile ? Inutile de se voiler la face.
Il revient à chacun de faire ses choix. Et il revient, aussi et surtout, à la collectivité de reconsidérer de fond en comble ses organisations et nos modes de vie.
La science constate, décrit, prévoit. Elle ne nous dit pas comment faire. À la politique, au sens le plus large et le plus noble du terme, de prendre le relai.
La science est tantôt glorifiée, tantôt décriée. Ne lui demandons pas ce qu’elle ne peut pas donner mais sachons tenir compte de ce qu’elle nous dit.
Le GIEC, dans son sixième rapport - et les cinq précédents n’ayant pas eu les effets escomptés - recommande la sobriété. La porte est étroite, le chemin long et difficile, alors ouvrons le débat, relevons le défi.
Alain Foucault
Professeur émérite du Muséum national d'histoire naturelle (Paris)

Après avoir publié, pour son sixième rapport, les travaux de ses deux premiers groupes de travail (voir sur le site de l’Afas les articles «Changement climatique, le GIEC persiste et signe» et «Le GIEC publie le deuxième tiers de son sixième rapport»), le GIEC (Groupe d’experts intergouvernemental sur l’évolution du climat) publie ceux de son troisième groupe de travail chargé de se pencher sur les actions pouvant être entreprises pour atténuer le réchauffement climatique.
Comme pour ses prédécesseurs, la lecture de ce rapport, gros en tout de 2913 pages, n’est pas chose aisée. Il est bien précédé d’un résumé pour les décideurs, mais qui ne peut rendre compte de la substance du texte complet, auquel il faut parfois se référer pour une bonne compréhension.
Une constatation donne le ton de ce rapport et sa leçon principale : nous avons déjà émis tellement de gaz à effet de serre qu’il ne nous en reste plus beaucoup à émettre si l’on veut rester dans les limites du raisonnable. Entre 2010 et 2019, les émissions cumulées de CO2 ont été de 410 Gt. Il ne faut pas ajouter plus du quart de cette quantité si l’on veut limiter le réchauffement à 1,5°C (500 Gt) et plus de deux fois cette valeur si l’on veut limiter le réchauffement à 2°C (1150 Gt).
Les auteurs ont examiné de très nombreux scénarios d’émission de gaz à effet de serre conduisant à des réchauffements plus ou moins rapides. Leurs conclusions sont inquiétantes : tous les scénarios qui limitent le réchauffement à 1,5°C et ceux qui le limitent à 2°C impliquent une réduction des émissions de gaz à effet de serre rapide et substantielle et, dans la plupart des cas, immédiate. Dans tous ces scénarios, les émissions mondiales doivent décroître au plus tard en 2025.
Les modélisations nous indiquent que ces réchauffements nous causeront des problèmes (voir sur le site de l’Afas l’article «Un nouveau rapport spécial du GIEC sur un réchauffement global possible de 1,5°C»). Mais si l’on se doute bien que ces problèmes intéresseront l’ensemble des activités humaines, les réactions de notre espèce sont bien difficiles à prévoir. Et cette absence de prévision sur ce qui va nous arriver explique peut-être notre manque d’empressement à prendre les mesures nécessaires.
Il est cependant urgent de prendre ces mesures pour limiter encore davantage et rapidement nos émissions de gaz à effet de serre. Pour cela, le rapport nous donne une quantité de solutions, touchant l’énergie, l’agriculture, les transports, les industries, etc.
Il ne reste plus qu’à les appliquer...